Selvreproduserende enzymer?

En kritikk av noen aktuelle evolusjonære 'opprinnelse-til-liv' modeller.
Evolusjonære 'opprinnelse-til-liv' teorier har mange hindringer å overvinne. (1,2,3). For å danne en selvreproduserende celle fra ikke-levende kjemikalier, krever generering av en stor mengde informasjon, eller spesifisert kompleksitet. En celle må være i stand til å utføre mange kjemiske reaksjoner i riktig rekkefølge, sted og grad, noe som krever en rekke spesifikke katalysatorer (enzymer). Det må også være i stand til å reprodusere den informasjonen som trengs for å produsere disse enzymene.
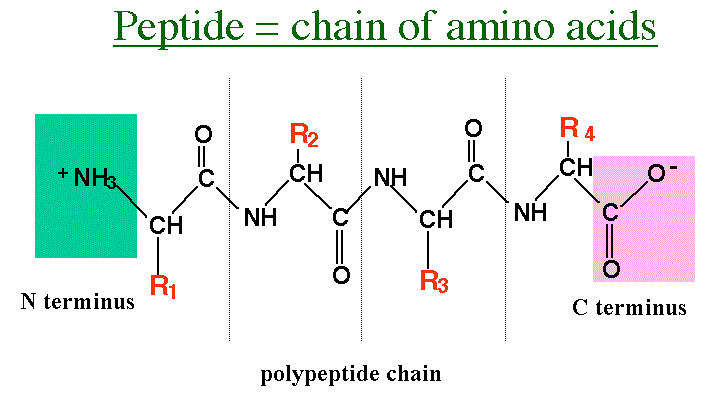
I alle kjente celler, er de spesifikke katalysatorene proteiner, mens informasjons lagring/henting og reproduksjons -oppgaver utføres av nukleinsyrer DNA og RNA. Proteiner er polymerer av aminosyrer, mens nukleinsyrer er polymerer av nukleotider. Nukleotider er i seg selv en blanding av et sukker (deoksyribose for DNA, ribose for RNA), en nitrogenholdig base og en fosfatgruppe.
Bilde 1. Livets opprinnelse -god fortjeneste i spekulasjon
Men DNA koder selv for proteiner, likevel kreves minst 50 proteiner for den nødvendige dekoding, og atter andre for replikering. Den kjente vitenskapsfilosof, den avdøde Sir Karl Popper, kommenterte:
Det som gjør livets opprinnelse og den genetiske koden til en urovekkende gåte er dette: den genetiske koden er uten biologisk funksjon med mindre det er oversatt; det vil si, hvis den ikke fører til syntese av proteiner som har struktur fastlagt av koden. Men ... maskiner ved hvilken cellen (i det minste den ikke-primitive celle, som er den eneste vi kjenner til) som oversetter kode, består av minst femti makromolekylære komponenter som selv er kodet i DNA. Dermed kan ikke koden oversettes uten ved hjelp av visse produkter av sin oversettelse. Dette utgjør en uforståelig sirkel; en virkelig ond sirkel, synes det som, for ethvert forsøk på å danne en modell eller teori, for dannelse av den genetiske koden.
Dermed kan vi bli møtt med muligheten for at livets opprinnelse (som opprinnelsen til fysikk) blir en ugjennomtrengelig barriere for vitenskap, og en gjenlevning av alle forsøk på å redusere biologi til kjemi og fysikk. (4)
Den åpenbare konklusjonen er at både DNA og proteiner må ha vært funksjonelle fra begynnelsen, ellers kunne ikke livet eksistere.
 RNA verden?
RNA verden?
Bilde 2. RNA
For å unngå denne konklusjonen, har noen evolusjonister laget en teori om at en type molekyl kunne utføre både katalytiske og reproduktive roller. En nylige oppdagelse av noen katalytiske funksjoner i RNA, har ført mange evolusjonister å postulere en "RNA verden". Tanken er at det første livet besto hovedsakelig av RNA, som ikke bare kunne reprodusere, men også utføre mange av funksjonene, nå utført av enzymer. Men denne modellen har flere tvilsomme postulater:
1. En pool av utelukkende "høyrehendt" ribose molekyler kunne produseres, adskilt fra et virvar av andre sukkerarter, og holde seg stabil lenge nok; basene kunne produseres i store mengder; og en høy konsentrasjon av fosfat (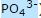 ) vil forbli i oppløsningen, i stedet for å utfelles.
) vil forbli i oppløsningen, i stedet for å utfelles.
2. Ribose kunne kombinere med baser og fosfat for å fremstille beta-D-ribonukleotider.
3. Disse beta-D-ribonukleotider kan spontant produsere RNA-polymerer av riktig slag.
4. At hvis slike polymerer dannes, kunne de kopiere seg selv.
5. At slike selvreproduserende RNA-molekyler ville ha alle funksjoner som trengs, for å opprettholde en organisme.
6. At en slikt RNA organisme kan gi opphav til en moderne organisme med protein katalysatorer, kodet på reproduksjonsmaterialet, og midler for å dekode dem.
Disse postulater er alle i strid med eksperimentelle bevis. (5) Det er ikke rart at en av de ledende forskerne i 'RNA World' modeller, Gerald Joyce, skrev:
Den mest rimelig antakelse er at livet ikke startet med RNA .... Overgangen til en RNA verden, som opprinnelsen til livet generelt, er fylt med usikkerhet og er plaget av mangel på eksperimentelle data. (6)
Et selvreproduserende Molekyl
En gruppe ledet av Julius Rebek, syntetiserte et molekyl som kalles Amino-Adenosin Triacid Ester (AATE), som i seg selv består av to komponenter, pentafluorfenyl ester og aminoadenosin. Når AATE molekyler er oppløst i kloroform med de to komponentene, fungerer AATE molekyler som maler for de to komponentene, til å forenes og danne nye AATE molekyler. (7) Det finnes en rekke grunner til at dette er irrelevant for en evolusjonær opprinnelse av livet:
1. Dette systemet medfører meget lite informasjon, i motsetning til selv den enkleste celle. Mycoplasma genitalia har den minste kjent genom av enhver levende organisme, som inneholder 482 gener som omfatter 580000 baser. (8) Denne organismen er en parasitt. En frittlevende organisme ville trenge mange flere gener.
2. Det nye AATE molekyl binder seg for sterkt til foreldrene, slik at ingen nye reaksjonspartnere kan komme inn og bli med, som Rebek selv innrømmer. (9)
3. Replikation skjedde bare i svært kunstige, unaturlige forhold. (10) En reaksjon i kloroform er irrelevant for levende organismer. Spesielt ville kloroform ikke hindre kondenseringsreaksjoner, slik vann gjør. De fleste polymerisasjonsreaksjoner i livet er kondenseringsreaksjoner, det vil si, de løser ut et lite molekyl som vann. Dersom det er mye vann rundt, som det er med alle levende ting, foretrekkes den reverserte reaksjonen, d.v.s. hydrolyse (oppløsning) av polymerer.
[For mer informasjon, se forfatters senere artikkel, Origin of Life: Polymeriserings Problemet].
4. Molekylet reproduseres for nøyaktig-det er ingen mulighet for neo-darwinistisk evolusjon ved mutasjon og naturlig utvalg. (11)
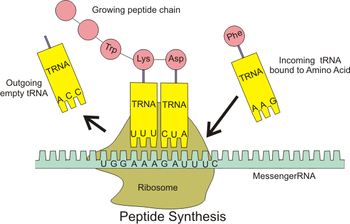
 Selvreproduserende Peptider?
Selvreproduserende Peptider?
Aminosyrer kan bli dannet, -med vanskelighet (12) i Miller-type forsøk hvor reduserende gasser blir antent, i motsetning til ribose og nitrogenholdige baser. Dermed undersøker noen evolusjonister 'proteiner først', snarere enn 'nukleinsyre-syre først', teorier om livets opprinnelse. Men proteiner har ikke noen analog til baseparing i nukleinsyrer. Så det var en overraskelse i august 1996, da noen aviser og tidsskrifter rapporterte et peptid som kan reprodusere seg selv. David Lee et al., rapporterte at et kort peptid avledet fra en del av et gjær-enzym kan katalysere sin egen dannelse. (1,3)
Lee et al. laget en 32-enhet lang, spiralformet peptid, basert på leucin, til gjær-transkripsjonsfaktor GCN4. De fant at den katalyserte sin egen syntese i en nøytral, fortynnet vannoppløsning av 15 og 17-enhets fragmenter. Dette var et genialt eksperiment, men det hjelper ikke den evolusjonære årsaken fordi:
1. Hvor ville den første 32-enhet lange kjede, av 100% venstrehendt aminosyreenheter, komme fra? Aminosyrer utformes ikke så lett som Lee et al. hevder. Dersom de dannes i det hele tatt, er de ekstremt fortynnede og urene, så vel som racemiske (50-50 blanding av venstre- og høyrehendte former). Slike aminosyrer polymerisere ikke spontant i vann.
2. Hvor ville en tilførsel av den tilsvarende 15 og 17-enhetskjedene komme fra? Ikke bare ville innvendingen ovenfor gjelde, men hvilken mekanisme er ment å produsere de riktige sekvensene? Selv om vi hadde en blanding av rette homokirale (alle de samme venstre-dreiende aminosyrer, riktige, er sjansen for å få en 15-enhet peptid en i 20 opphøyd i 15 (= 1 : 3 x 10 opphøyd i 19). Dersom det ikke er nødvendig for å få sekvensene helt riktig, så ville det bety at "replikasjon" ikke er spesifikk, og derfor vil tillate mange feil.
3. De 15 og 17-enhets peptider må aktiveres, fordi kondensasjon av vanlige aminosyrer ikke er spontane i vann. Lee et al., benyttet en tiobenzyl-ester-derivat av en peptid. Som de sier, omgår dette også potensielle sidereaksjoner. Den hypotetiske ursuppen ville ikke ha hatt intelligente kjemikere som la til riktige kjemikalier, for å hindre feil reaksjoner!
Bilde 3. Peptide -kjede av aminosyrer
4. Den spesielle 32-enhets kjeden var en en alfa-heliks, der hydrogenbindinger mellom forskjellige aminosyrere-rester, får kjedet til å helicidere (helicize). Denne felles struktur er mer sannsynlig å være i stand til å fungere som en mal i henhold til kunstige betingelser. Lee et al. hevder at beta-lag (sheets), som også er avhengige av hydrogenbinding, også kan være i stand til å fungere som maler. Dette synes sannsynlig. Alfa-helikser og beta -sheets er kjent som den sekundære struktur til proteinet. (14)
Den nøyaktige måte som proteinet folder seg, kalles den tertiære struktur, og dette bestemmer dens spesifikke egenskaper. . Selv om Lee et al sier:
"vi foreslår muligheten for protein selv-replikasjon, hvor den katalyserende aktivitet til proteinet kan bli konservert", så presenterer de ingen eksperimentelle bevis.
 Kompleksitets Teori
Kompleksitets Teori
Dette er blitt fremmet av Stuart Kauffman. (15) Den hevder at et stort antall samvirkende komponenter, spontant organiserer seg i ordnede mønstre. Noen ganger kan en liten forstyrrelse av et system føre til at det skifter fra et mønster til et annet. Kauffman foreslår at hans idé kunne redegjøre for livets opprinnelse, kroppsformer og selv kulturelle mønstre og økonomi. Kompleksitet teoretikere peker på datasimuleringer av mønstre av musling- skjell og andre former som finnes i naturen.
Men dette har liten relevans til den virkelige verden av kjemikalier. .. Levende celler har molekylære maskineri for å kanalisere kjemien i riktig retning og mengder. Hvis et muslingskall-mønster på dataskjermen ble forstørret, ville det ikke være noen spor av celler med flimmerhår, mitokondrier, DNA, etc. (16)
Det er rart at selv de fleste deler av det evolusjonære etablissement er skeptiske til kompleksitetsteori. Omslaget på juni-1995 utgaven av Scientific American spurte "Er kompleksitetsteori et falskneri? '. Dette utgaven inneholdt en artikkel som het "Fra kompleksitet til rådvillhet ', som sa: "Kunstig liv, et stort delfelt av kompleksitets-studier, er "fakta-fri vitenskap", ifølge en kritiker. Men det utmerket seg ved å generere datagrafikk. (17)
Bilde 4. Vanskelig å forstå det enkleste
Relaterte Artikler
Referanser
 1. En utmerket og up-to-date oppsummering er Aw, S.E., 1996. The Origin of Life: A Critique of Current Scientific Models. Journal of Creation 10(3):300-314 (Download PDF file).
1. En utmerket og up-to-date oppsummering er Aw, S.E., 1996. The Origin of Life: A Critique of Current Scientific Models. Journal of Creation 10(3):300-314 (Download PDF file).
2. Thaxton, C. B., Bradley, W. L. & Olsen, R. L., 1984. The Mystery of Life’s Origin, Philosophical Library Inc., New York. This is a very thorough treatment of chemical and thermodynamic objections to evolutionary origin-of-life theories.
3. Kompleksiteten i cellens maskineri er godt illustrert av Behe, M.J., 1996. Darwin’s Black Box: The Biochemical Challenge to Evolution, The Free Press, New York.
4. Popper, K.R., 1974. Scientific Reduction and the Essential Incompleteness of All Science. In Ayala, F. and Dobzhansky, T., eds., Studies in the Philosophy of Biology, University of California Press, Berkeley, p. 270.
5. Se Mills, G.C. and Kenyon, D.H., 1996. The RNA World: A Critique. Origins and Design, 17(1): 9-16.
6. Joyce, G. F., 1989. RNA evolution and the origins of life. Nature, 338:217-224.
7. Tjivikua, T., Ballester, P. and Rebek, J., Jr., 1990. A Self-Replicating System. J. Amer. Chem. Soc., 112(3):1249-50.
Bilde 5. Produksjon av peptider- i virkelighetens verden
8. Fraser, C.M., et al. 1995. The minimal gene complement of Mycoplasma genitalium, Science, 270(5235):397-403; Perspective by A. Goffeau. Life with 482 genes. Same issue, pp. 445-6.
9. Amato, I., 1990. Making molecules that copy themselves. Science News, 137(5):69.
10. Horgan, J., 1991. In the Beginning. Scientific American, 264(2):100-109, reporting a comment by Gerald Joyce on p. 104.
11. Horgan, ref. 10.
12. Se Thaxton et al., Ref. 2.
13. Lee, D.H., Granja, J.R., Martinez, J.A., Severin, K. And Ghadiri, M.R., 1996. A self-replicating peptide. Nature, 382:525-528. See also Kauffman, S. 1996. Even peptides do it. Nature, 382:496-497, for a perspective of the leading complexity theorist on the paper of Lee et al.
14. Behe, Ref. 3, pp. 259-265 explains such terms as a-helices and b-sheets, secondary and tertiary structures.
15. Kauffman, S. A., 1993. The Origins of Order, Oxford University Press, Oxford, UK.
16. Behe, Ref. 3, pp. 189-192 raises such objections.
17. Horgan, J., 1995. From Complexity to Perplexity. Scientific American, 272(6):74-79. The critic was the well-known evolutionary theorist John Maynard Smith, see p. 77.
Stoffutvalg og bilder ved Asbjørn E. Lund